文/阿瑶说
编辑/阿瑶说


前言
脊椎动物在进化过程中出现了磷酸钙基生物矿化组织,这使得它们能够发展各种防护盔甲、牙齿和内部骨骼。生物矿化骨骼最早出现在大约5亿年前的水生脊椎动物中,作为覆盖基底无颌鱼类身体的真皮外骨骼。
随着进化的进行,这种外骨骼盔甲逐渐减少和改造,而矿化的内部骨骼变得更加显著。因此,在现存的脊椎动物中,骨骼要么完全由骨质构成(如大多数脊椎动物),要么由矿化项圈加固的软骨核心组成(如鲨鱼和鳐鱼)。

无论主要骨骼组织类型是矿化软骨还是骨骼,所有脊椎动物的矿化组织都由水、非胶原蛋白和碳酸磷灰石生物矿化的胶原纤维组成。
因此,脊椎动物骨骼组织中观察到的材料特性范围并不是由化学成分的变化决定的,而是通过构建块(蛋白质、生物矿物质和水)的比例变化以及组织结构来确定。例如,哺乳动物骨骼的矿物质含量从约50%到接近100%不等,导致刚度和韧性的巨大变化。

哺乳动物骨骼的多样性使其成为研究组成和结构对生物矿化组织材料特性影响的有用系统。存在具有极高或极低矿物质密度的骨组织(如耳骨和鹿角骨),具有相似体积矿物密度但具有显著不同的超微结构或中观结构的骨组织(如骨和纤维层以及皮质和松质骨),以及骨组织中存在局部差异的例子(如骨和骨皮质骨与纤维层和骨区)。
对这些组织的研究有助于理解骨骼结构和组成复杂性。

然而,这些研究主要集中在少数哺乳动物物种上,忽略了其他脊椎动物在生态、骨骼和骨骼组织类型方面的广泛多样性。
为了更全面地了解脊椎动物骨骼组织在结构、组成和力学方面的真正变异范围,需要广泛研究多样的脊椎动物。
通过研究哺乳动物骨骼的进化替代物,可以扩展和重新构建对脊椎动物骨骼组织形式与功能关系以及驱动其进化的因素的理解。

特别是,鱼类骨骼组织——软骨和硬骨鱼类(即软骨鱼和硬骨鱼)——为定义和理解脊椎动物骨骼组织中形式与功能关系的参数空间提供了独特的机会。软骨鱼和硬骨鱼占据了广阔的栖息地和生态位,表现出广泛的形态多样性,它们占了所有脊椎动物物种的50%以上。
此外,它们的骨骼由与哺乳动物骨骼相同的基本成分构成,但具有特定的中观结构排列,导致独特的力学性能。尽管这些鱼类群作为骨骼生物学模型具有优势,但对鱼类骨骼组织中结构与功能关系的研究仍处于初步阶段。
本文摘录了一章的内容,该章节回顾了硬骨鱼和软骨鱼(即鲨鱼和鳐鱼)中镶嵌软骨组织的多个层次的结构与功能关系的现有知识。特别关注了镶嵌软骨的超高分辨率超微结构数据,并讨论了这些特征与现有力学数据之间的潜在联系。
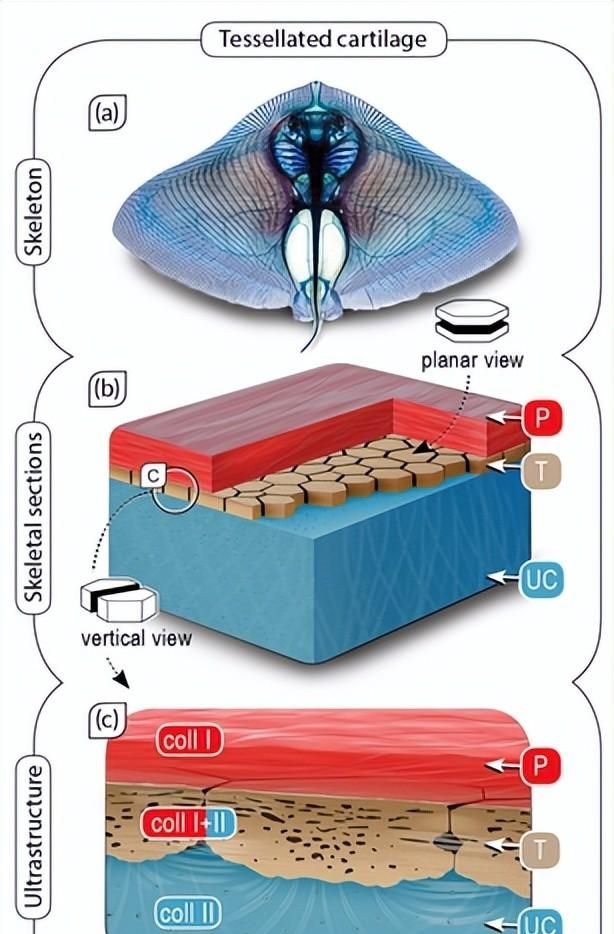
鲨鱼和鳐鱼镶嵌软骨
结构
鲨鱼和鳐鱼通常被称为“软骨鱼”,指的是这些鱼的骨骼与绝大多数其他脊椎动物的骨架的不同之处。像大多数脊椎动物一样,鲨鱼和鳐鱼会发育出胚胎未矿化的软骨骨骼;尽管如此,在个体发育过程中,这从未被骨骼取代,而是在其一生中保持大部分软骨。骨骼和非矿化软骨显然是用于构建骨骼的完全不同的材料。
在(a)组织组织(骨骼和软骨分别在I型和2型胶原蛋白上形成图案),(b)材料特性(在大多数生理负荷状态下,骨骼比软骨硬约10000倍)和(c)对组织损伤的反应(与骨骼不同,软骨的血管和神经供应有限, 大多数脊椎动物软骨在受伤后无法自我修复)。
鲨鱼和鳐鱼骨骼的独特结构图案使软骨能够执行许多与其他98%脊椎动物骨骼相同的机械作用:大多数软骨内骨骼被一层薄薄的矿化多边形瓷砖覆盖,称为tesserae,通常宽和深数百微米。
这种生物矿化组织的镶嵌外壳夹在骨骼的未矿化软骨核心和包裹每个骨骼元素的外部纤维周骨之间,形成分层的纤维矿物复合材料。这种独特的内骨骼瓷砖通常仅占每个骨骼元素体积的30%或更少,但似乎是软骨鱼类的重要进化创新。
Tesserae已经表征了超过400亿年的软骨骨骼,对鲨鱼和鳐鱼骨骼生物学至关重要。Tesserae不仅允许骨骼软骨核心及其生物矿化层的生长 – 通过在tesserae边缘沉积新的生物矿物,一种生长机制,具有连续的生物矿化地壳,无法重塑 – 但也为骨骼元素提供了刚度(见“力学”部分)。
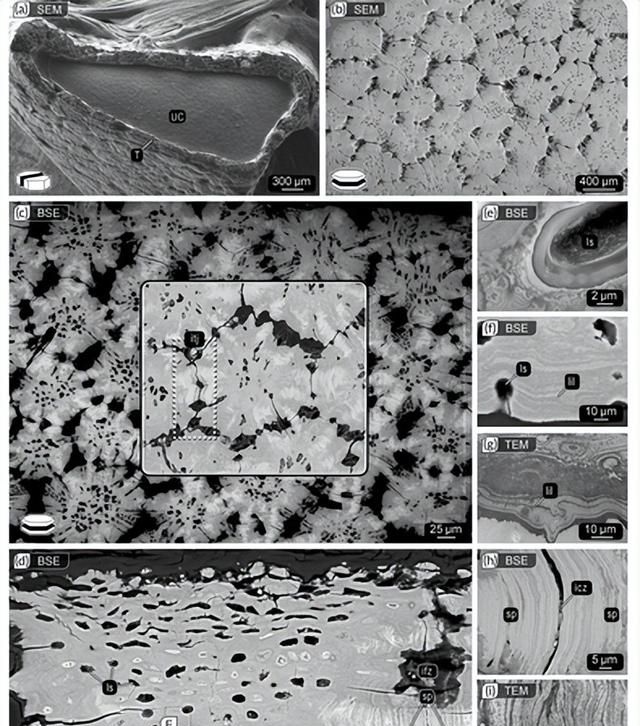
就镶嵌内骨骼的一般发育和超微结构而言,狗鱼Scyliorhynus canicula和圆形黄貂鱼Urobatis halleri可能是研究得最好的软骨鱼类。
在发育过程中,tesserae首先出现在胚胎骨骼中,作为软骨钙化的孤立血小板,嵌入未矿化的软骨并被其分离。单个瓷砖通过生物矿物在其现有表面上的吸积而生长,这一过程反映在生物矿化组织中不同矿物密度的周期性同心层(Liesegang 线)中。
这种增生生长最终使年轻的镶嵌物在它们的侧缘相互接触。一旦发生这种情况,在两个相邻镶嵌物的直接接触区域就会形成明显的高矿物密度特征,称为“辐条”。

前面描述的镶嵌软骨的一般结构特征和组织排列似乎在很大程度上是鲨鱼和鳐鱼的普遍特征。然而,高分辨率电子显微镜和同步加速器断层扫描数据表明,镶嵌的形状和结构在个体内部(例如,骨骼的不同区域之间)和物种之间都可能有所不同,这进一步表明镶嵌物之间的相互作用在功能上很重要。
例如,不同鲨鱼和鳐鱼物种的镶嵌物在尺寸(宽度和厚度从 <100 μm 到近 1 毫米)和形状上差异很大,从圆盘状板到星形形式再到长方体块。
镶嵌物的形状似乎也根据它们在骨骼元素上的位置而变化,并根据它们覆盖的骨骼元素(例如,在某些鲨鱼的嘴软骨和某些鳐鱼的颌骨的特定区域中始终如一地长方体。这些观察结果强烈表明,镶嵌物的形状、它们的关节以及镶嵌垫和整个骨骼元件的有效机械行为之间存在形式-功能关系,但这些联系才刚刚开始建立。
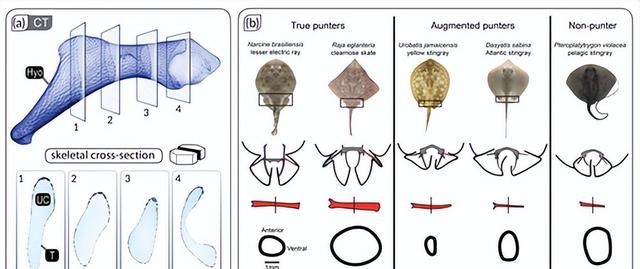
力学
一般来说,镶嵌软骨力学的研究远远落后于骨骼解剖学和镶嵌超微结构的研究。尽管如此,几项工作的结果结合在一起,开始描绘出软骨软骨的结构和机械性能如何相互关联,以及tesserae如何在根据特定的生态角色和高承重活动定制骨骼特性方面发挥重要作用。
要了解镶嵌软骨(一种具有相对离散材料相的复合材料)的整体机械性能,就需要表征主要组织成分的特性:未钙化的软骨、未矿化的关节纤维和矿化组织。

然而,由于镶嵌软骨的精细3D结构排列以及需要模拟生理条件(例如水合和负荷率)的测试条件,捕获体内或原位特性变得复杂。有限的可用证据表明,软骨鱼未钙化软骨具有类似于哺乳动物透明软骨的蛋白多糖和胶原蛋白含量,并建议在类似的负载速率下,它至少可以像刚性一样硬几个数量级。
这些特性显然取决于所测试的物种和骨骼元素;需要更严格的研究来了解机械性能与成分、加载速率和系统发育之间的关系。
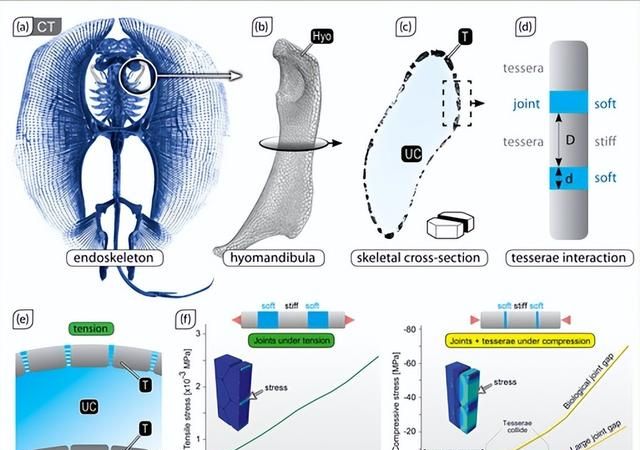
虽然用坚硬的连续壳覆盖软骨样凝胶有望增加刚度但降低复合材料的柔韧性,但有一些迹象表明,来自镶嵌软骨但也制造阵列,具有相互作用瓷砖的镶嵌壳可以是“两全其美”的配置,最大限度地提高两种组织相的理想特性。
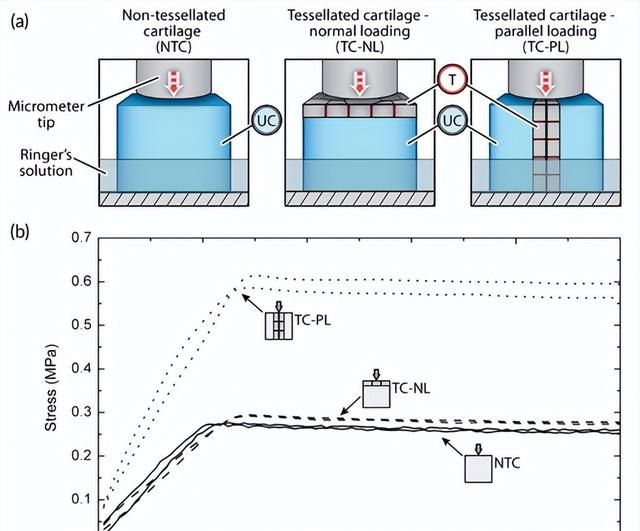
Fahle和Thomason表明,与胚胎(非镶嵌)小斑点猫鲨(S. canicula)相比,成年个体的颌软骨具有更高的抑制机械能的能力,但也更硬。刚度的很大一部分肯定是由于成年动物的镶嵌层。从生物学的角度来看,这种特性的变化允许成年人比新生儿消耗更硬的猎物,但对于工程考虑也特别有趣,因为刚度和阻尼通常在人造材料中呈负相关。
镶嵌层相对于加载方向的排列在软骨骨骼组织力学中起着相当大的作用。来自蓝鲨(Prionace glauca)的镶嵌软骨立方体垂直加载到镶嵌垫(在应力松弛实验中)的行为与非镶嵌立方体相似,比镶嵌立方体软~45倍,其中负载在平面上施加与镶嵌垫。
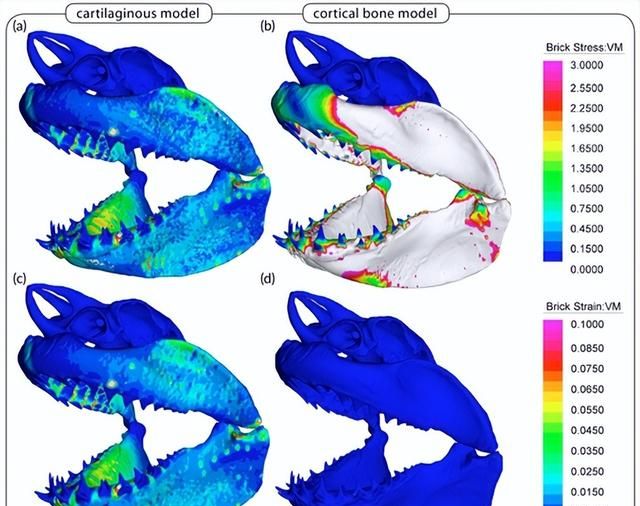
这些结果得到了对两条大型鲨鱼的水合颌骨样品进行的压痕实验的支持。纳米压痕实验通常涉及将非常小的硬尖端(例如,尖端半径为数百纳米)推入材料中,以检查非常小尺度下的硬度和弹性模量。
尽管如此,作为费拉拉等人使用的压头。相对较大(100μm),并且接近一些镶嵌物的尺寸,据信他们的数据更能代表复合材料(例如,tesserae及其周围的软组织)的特性,因为它们报告的值比tesserae本身要软得多。
事实上,最近的研究表明,镶嵌中的一些细尺度结构特征(例如,辐条中的高矿物密度薄片)表现出超过哺乳动物骨骼和钙化软骨的刚度和硬度值,并且具有更类似于牙齿材料的特性,需要深入研究局部属性和涌现骨架属性之间的关系,以更好地了解不同层次的结构层次结构在骨架性能中的作用。
前面描述的广义镶嵌软骨解剖结构的变化,当在动物生态学的背景下解释时,也提供了关于体内骨骼性能以及组织功能极限的观点。
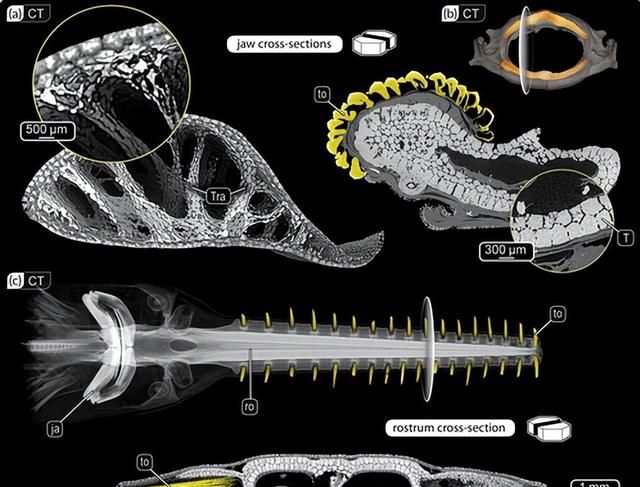
例如,除了外镶嵌层外,许多蝙蝠鱼(鳐鱼和亲戚)的下颌还含有空心镶嵌支柱(小梁),通常直径数百微米,横跨颌骨的未钙化软骨核心。这些似乎沿着主要载荷线排列,通常在颌骨或颌关节的狭窄区域,因此在结构和功能上与四足动物中发现的小梁骨收敛。
小梁对加强镶嵌软骨的重要性通过在进食期间经历高骨骼负荷的物种的颌骨中高密度的小梁得到强调,例如较小的电射线,它使用爆炸性的下颌突起来取回埋藏的猎物,或者黄貂鱼和吉他鱼,它们使用高咬合力来粉碎硬壳软体动物。
结论
尽管哺乳动物骨骼通常被认为是典型的脊椎动物骨骼组织,但海洋中存在许多其他鱼类,其中软骨鱼的镶嵌软骨是一种自然界的骨骼设计替代方案,为更广泛地研究生物矿化材料中形式和功能之间的相互作用提供了宝贵的资源。
镶嵌软骨还提供了大量未开发的机会,可以探索具有其他相似成分组织中交替结构的机械效应(例如,镶嵌软骨中的胶原蛋白和碳酸磷灰石成分类似于骨组织)。显然,镶嵌软骨中软组织和硬组织的不同几何排列具有深远的机械效应,例如刚度和阻尼/柔韧性的令人印象深刻的组合。
此外,由于该组织的某些结构方面可以通过简单的几何形状近似(例如,镶嵌瓷砖的多边形),因此该系统特别适合对生物设计的参数空间进行建模探索。例如,可以研究为什么某些形态已经进化并且与某些加载状态相关联,而其他形态则没有。考虑到这些因素,软骨鱼的镶嵌软骨成为各种学科的有用模型,特别适用于研究生态学与生物材料之间的联系(鲨鱼和鳐鱼表现出广泛的饮食多样性),结构瓷砖的生物学优势和机械效应,以及分层组织复合材料的设计和行为。
参考文献:
Simon J. Penn, Steven C. Sweetman, Microvertebrate-rich gutter casts from the basal WessexFormation (Wealden Group, Lower Cretaceous) of Dungy Head, Dorset: Insights into thepalaeoecology and palaeoenvironment of a non-marine wetland, Cretaceous Research,10.1016/j.cretres.2022.105397,143,(105397),(2023)
Merle Greif, Humberto G. Ferron, Christian Klug, A new Meckel’s cartilage from the DevonianHangenberg black shale in Morocco and its position in chondrichthyan jaw morphospace, Peer10.7717/peerj.14418,10,(e14418),(2022).
Manuel Andreas Staggl, Daniel Abed-Navandi, jurgen Kriwet, Cranial morphology of theorectolobiform shark, Chiloscyllium punctatum Muller & Henle, 1838, Vertebrate Zoology.10.3897/vz.72.e84732,72,(311-370),(2022).
Eduardo Villalobos-Segura, Giuseppe Marrama, Giorgio Carnevale, Kerin M. Claeson, Charlie J.Underwood, Gavin J. P. Naylor, Jurgen Kriwet, The Phylogeny of Rays and skates (Chondrichthyes:Elasmobranchi) Based on Morphologica Characters Revisited, Diversity, 10.3390/d14060456. 14. 6(456),(2022).
本文来自投稿,作者:时遇,不代表食养源立场,如若转载,请注明出处:https://www.xiayuan17.com.cn/yszs/98953.html